This chapter in pdf-format: www.researchgate.net/publication/321137184
This chapter in pdf-format: www.researchgate.net/publication/321137184
8
Source-Field Models
The simplest source configuration is the point source or monopole. If we consider a point current source of magnitude I0 lying in a uniform conducting medium of infinite extent and conductivity σ, then current flow lines must be uniform and directed radially. As a consequence, for a concentric spherical surface of arbitrary radius r, the current density J crossing this surface must be uniform and will equal I0 divided by the total surface area. That is
 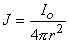 | (8.1) |
since the total current is conserved. Because the current is everywhere in the radial direction, the current density expressed as a vector is
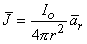 | (8.2) |
where 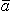 r = unit vector in the radial direction, where the origin is at the point source.
r = unit vector in the radial direction, where the origin is at the point source.
Associated with the current flow field defined by Equation 8.2 is a scalar potential field Φ. Since the field is everywhere radial, there should be no variation of potential along a transverse direction, namely that on which r is a constant. Consequently, we expect isopotential surfaces to be a series of concentric spheres surrounding the point source with diminishing potentials for increasing values of r. In a formal sense, it is known from field theory that the electric field 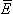 is related to a scalar potential Φ by
is related to a scalar potential Φ by
= -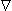 Φ Φ | (8.3) |
From Ohm's law it follows that
 = σ = σ | (8.4) |
Applying Equations 8.3 and 8.4 to 8.2 results in
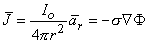 | (8.5) |
To satisfy Equation 8.5, only the component of Φ in the direction of r can arise. This leads to
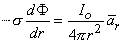 | (8.6) |
and integration with respect to r leaves us with
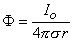 | (8.7) |
As suspected above, Φ is a constant on surfaces where r is constant (i.e., concentric spheres). Normally the potential for r 
 is set to zero, which accounts for having chosen the constant of integration in Equation 8.7 equal to zero. We note from Equation 8.7 that equipotential surfaces are indeed concentric spheres and that the potential magnitude is inversely proportional to the radius (with the origin at the monopole source).
It is not always convenient to place the coordinate system origin at the point source (e.g., when considering several such sources). In this case it is desirable to distinguish the coordinates of the point source(s) from that of the field point, and we do this by using primes for the field point coordinates. Equation 8.7 then applies with, r given by
is set to zero, which accounts for having chosen the constant of integration in Equation 8.7 equal to zero. We note from Equation 8.7 that equipotential surfaces are indeed concentric spheres and that the potential magnitude is inversely proportional to the radius (with the origin at the monopole source).
It is not always convenient to place the coordinate system origin at the point source (e.g., when considering several such sources). In this case it is desirable to distinguish the coordinates of the point source(s) from that of the field point, and we do this by using primes for the field point coordinates. Equation 8.7 then applies with, r given by
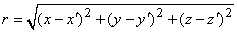 | (8.8) |
where each monopole is located at (x, y, z) while the field point is at (x', y', z').
The field described by Equation 8.7 for a point current source is identical to the electrostatic field from a point charge, provided that I0 is replaced by Q0 (the charge magnitude), σ is replaced by ε (the permittivity), and replaced by . This result is not surprising since if the aforementioned exchanges are made, the governing equations for current flow convert exactly into those for electrostatics. This means that simply by interchanging symbols, solutions to problems in electrostatics can be converted into solutions to equivalent problems in current flow (and vice versa).
The aforementioned is an example of duality. It can be a useful tool when there is an extensive literature already in existence. Sometimes there may be a limitation in physically realizing a condition in one or the other dual systems. For example, one can have zero conductivity, but the permittivity can never be less than that of vacuum. Also, while one can have a point charge, one cannot actually have a physical point source.
The reader may wonder why there is an interest in a point current source when such is not physically obtainable. One reason is that in a limited region, the fields may behave as if they arise from such a source (we say that the source is equivalent). Second, one can actually have two point sources of opposite polarity, in which case the field of interest can be found by the superposition of point source fields. In fact, this very situation is examined in the next section.
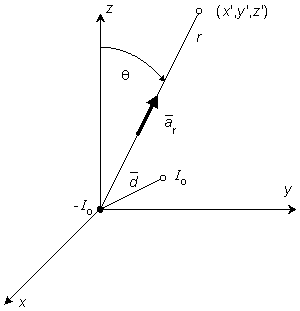
In bioelectricity one can never have a single isolated monopole current source because of the need to conserve charge. But collections of positive and negative monopole sources are physically realizable if the total sum is zero. The simplest collection, and one that reflects a fundamental bioelectric source, is the dipole. The dipole consists of two monopoles of opposite sign but equal strength I0 (often termed source and sink) separated by a very small distance, d. In fact, the strict definition requires d 0, I0 with p = I0d remaining finite in the limit. The quantity p is the dipole moment or dipole magnitude. The dipole is a vector whose direction is defined from the negative point source to the positive. In fact, if 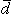 is the displacement from negative to positive point source and d a unit vector in that direction, then
is the displacement from negative to positive point source and d a unit vector in that direction, then
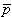 = Io = Io dd = Io = Io dd | (8.9) |
where = the dipole vector.
A dipole of arbitrary orientation is illustrated in Figure 8.1, where the coordinate system origin is placed at the negative pole. If the positive pole were also at the origin, the sources would cancel each other and their field would be zero. Consequently, the field arising from the displacement of the positive pole from the origin to its actual position (shown in Figure 8.1) is, in fact, the dipole field. But this can be found by examining the expression describing the potential of the positive monopole and evaluating the change in potential brought about by moving the monopole from the origin to its dipole position. And this, in turn, can be approximated from the first derivative of the monopole's potential field with respect to the source coordinates evaluated at the origin (as in a Taylor series representation). Specifically, to obtain the dipole field, a derivative of Φ (as given in Equation 8.7) is taken with respect to the direction (a directional derivative) and then multiplied by the magnitude of d. Thus, denoting the dipole field Φd, and based on Equation 8.7, we have
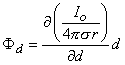 | (8.10) |
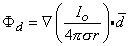 | (8.11) |
and, finally since I0d = p
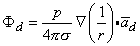 | (8.12) |
The accuracy of Equation 8.10 improves as d 0, and in fact, p (as noted earlier) is normally defined in the limit that d 0, I , such that the product I0d is finite and is equal to p. Consequently, Equation 8.12 is a rigorous (exact) expression for a mathematically defined dipole.
If the coordinate axes are oriented so that the dipole is directed along z- (the polar) axis and the dipole is placed at the origin, then carrying out the gradient operation in Equation 8.12 and noting that
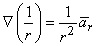 | (8.13) |
where r is oriented from the source point to field point, we obtain for the field of a dipole
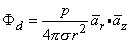 | (8.14) |
and
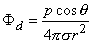 | (8.15) |
In Equation 8.15 the angle θ is the polar (colatitude) angle. The above expressions can be confirmed by noting that the gradient operator (in Equation 8.13) acts on the source (unprimed) coordinates in Equation 8.8.
A comparison of the dipole field to a monopole field, by contrasting Equation 8.15 with Equation 8.7, shows that the dipole field varies as (1/r)2 whereas the monopole field varies as (1/r). In addition, the dipole equipotential surfaces are not concentric spheres but, rather, are more complicated, because of the factor cosθ. The maximum dipole potential, for a given value of r, is on the polar axis (z axis).
Figure 8.2 illustrates a long, thin excitable fiber lying in a uniform conducting medium of conductivity σo and of unlimited extent. If we assume the existence of a propagating nerve impulse, then the activation currents are associated with a transmembrane current distribution im(x). Since the fiber is very thin and there is axial symmetry, we can describe the transmembrane current as a function of the axial variable x only. Thus the source description is one-dimensional. The dimension of im(x) is current per unit length. A small element of current im(x)dx can, therefore, be considered to behave like a point current source (a monopole) within the extracellular medium. Consequently, from Equation 8.7, we have
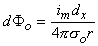 | (8.16) |
where r is given by Equation 8.8, Φo is the potential field and σo is the conductivity outside the fiber (i.e.,extracellular conductivity). Integration over the fiber (i.e., with respect to x) gives the total field as
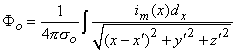 | (8.17) |
where the source is assumed to lie on the fiber axis, at (x, 0, 0), and the (fixed) field point is at (x', y', z').
We may apply the equations derived in Chapter 3, Section 3.4.2, to the fiber in Figure 8.2. We may approximate that the resistance of the interstitial medium ro  0 and that similarly the potential in the interstitial medium Φo 0. Using these approximations and Equation 3.42 and noting that Φi - Φo Vm we obtain
0 and that similarly the potential in the interstitial medium Φo 0. Using these approximations and Equation 3.42 and noting that Φi - Φo Vm we obtain
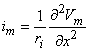 | (8.18) |
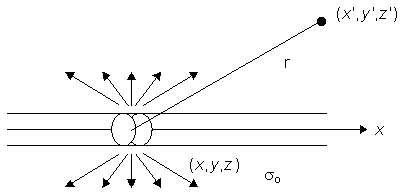
so that Equation 8.17 may be written
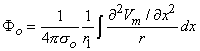 | (8.19) |
In Equation 8.19, r is given by
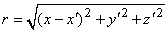 | (8.20) |
Using the cylindrical resistance formula for ri = 1/(πa2σi) based on a conductivity σi inside the cell, converts Equation 8.19 into
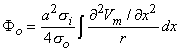 | (8.21) |
where a = the fiber radius.
The reader will note that initially Φo was set equal to zero and now we have found a solution for Φo which, of course, is not zero. The underlying explanation of this apparent paradox is that Φo was ignored in deriving Equation 8.18 in comparison with Φi. Since the latter is perhaps 100 times larger, dropping Φo at that point should have negligible consequences. The interested reader can pursue the matter by introducing the value Φo found in Equation 8.21 into the rigorous version of Equation 8.18, namely
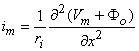 | (8.18b) |
and then recalculating Φo. This will produce an improved Φo. In fact, this iterative procedure can be repeated until a desired degree of convergence results. Such a procedure is followed in Henriquez and Plonsey (1988), and is seen to converge very rapidly, demonstrating that for typical physiological situations the first-order approximation (given by Equation 8.21) is entirely satisfactory.
Equation 8.21 may be integrated by parts. Since at the boundaries of the spatial activation, resting conditions are present,  Vm/x = 0 and the integrated term drops out. Accordingly, we are left with
Vm/x = 0 and the integrated term drops out. Accordingly, we are left with
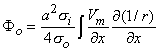 | (8.22) |
or
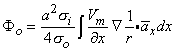 | (8.23) |
where x = unit vector in the x-direction.
Since both Equations 8.23 and 8.21 are mathematically the same, they necessarily evaluate the same field Φo. The physical interpretation of these expressions is that in Equation 8.21 the source is a (monopole) current density that lies on the axis, whereas in Equation 8.23 it is an axial dipole also lying along the axis. These are, of course, equivalent sources. Which source is prefeable to use depends on the shape of Vm(x); this will be illustrated in the following sections.
There are two approximations that underlie Equation 8.17 and that should be kept in mind. First, the configuration of the current element is approximated as a point source, but the current actually emerges from the membrane surface rather than a point (see Figure 8.2), and an axial segment could be characterized as a "ring source." For thin fibers this should be an acceptable simplification. Second, the field expression in Equation 8.17 is strictly for a point source in an unbounded space, whereas in reality the space is occluded by the fiber itself. This approximation is normally satisfactory. If, however, the extracellular space is itself limited, then the fiber probably cannot be ignored and the actual boundary value problem must be solved (Rosenfalck, 1969).
The unbounded extracellular space is important to justify not only the use of the "free-space" point source field of Equation 8.7 but also the linear core-conductor expression of Equation 8.18, which is based on the assumption that ro 0 and Φi - Φo Vm. For the isolated fiber of "small" radius, Equations 8.21 and 8.23 appear to be well justified (Trayanova, Henriquez, and Plonsey, 1990).
2Vm/x2)dx behaves like a point current source. Accordingly, the term (πa2σi2Vm/x2) has the dimensions of current per unit length. This is a function of x, in general; the variation with x constitutes a description of the source density strength. Since, in fact, the source is considered as lying on the axis, one can interpret the term (πa2σi2Vm/x2) as a line source density. This is a conceptual as well as a quantitative view of the origins of the volume conductor field (arising from the action potential described by Vm(x)).
Alternatively, one can group the terms in Equation 8.21 as
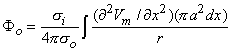 | (8.24) |
and σi2Vm/x2 now has the dimensions of a volume source density (flow source density) since πa2dx is a volume element. In fact, the interpretation of Equation 8.24 is that the source fills the intracellular fiber volume, where each source element is a disk of volume πa2dx. The source density is uniform over any disk cross section.
Of course, neither the volume nor line source is physically real. These sources are therefore designated as equivalent sources. That is, they are equivalent to the real sources in that the extracellular fields calculated from them are correct. For the calculation of intracellular fields the true sources (or some other equivalent source) would be required. We return to this topic in a subsequent section of this chapter.
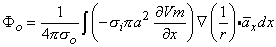 | (8.25) |
One can now identify a dipole element as (-σiπa2Vm/x)dxx. The dipole is oriented in the positive x-direction, and the line dipole density is (-σiπa2Vm/x).
Alternatively, the dipole source can be grouped as (-σiVm/x)x(πa2dx), which identifies (-σiVm/x) as a volume dipole density; this fills the intracellular space of the fiber, is oriented in the x-direction, and is uniform in any cross section. Hence, a dipole element also can be thought of as a disk of volume (πa2dx) with the vector magnitude of (-πa2dxσiVm/x)x.
2Vm/x2, which is shown schematically in Figure 8.3. Note that positive sources lie in the region x1 < x < x2 and x3 < x < x4, where the function 2Vm/x2 > 0, whereas negative sources lie in the region x2 < x < x3, where 2Vm/x2 < 0. The sum of the positive sources equals the net negative source. That the field outside the cell, generated by this source, is observed to be triphasic (two regions of one polarity separated by a region of opposite polarity) is not surprising in view of the triphasic source distribution.
When the distance to the field point is large compared to the axial extent of each positive or negative source region, then each such source can be approximated by a single (lumped) monopole at the "center of gravity" of the respective source distribution. This is illustrated in Figure 8.3. The resultant model is referred to as a tripole source model (since it consists of three monopoles). Intuitively we expect it to be valid, provided a representative distance from each source distribution to the field point ri satisfies
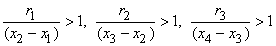 | (8.26) |
where r1, r2, and r3 as well as x1, x2, and x3 are as described in Figure 8.3. On the basis of Equation 8.24, we can express the tripole field as
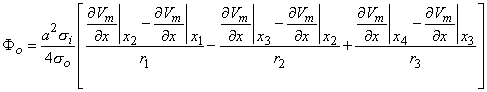 | (8.27) |
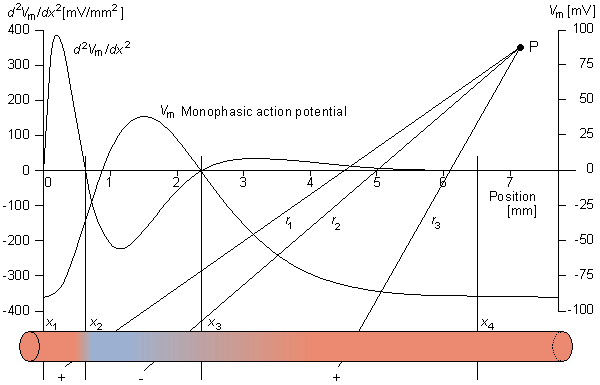
2Vm/x2 are shown. As explained in the text, the volume source density is proportional to 2Vm/x2. Consequently, positive sources lie in the region x1 < x < x2 and x3 < x < x4 while negative sources are present in the region x2 < x < x3. The sources within the fiber are illustrated below. When the extent of each source distribution is small compared to the distance to the field, each distribution can be summed into the lumped source as shown. The distances r1, r2, and r3 are from each lumped source to the distant field point P.
The expression for volume dipole density in Section 8.3.2 was given as (-σiVm /x), but this was derived for an isolated fiber. For the fiber bundle this can be shown to be (-σiCVm /x) (Plonsey and Barr, 1987), where C is a constant that depends on conductivities inside and outside the cell and the fiber bundle geometry. Its value is normally 0.4.
Figure 8.4 illustrates propagation of the rising phase of a cardiac action potential along a uniform bundle of fibers. In this figure the leading and trailing edges of the active region (where Vm /x 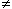 0) are assumed to be planar. All fibers in the bundle are assumed to be parallel and carrying similar action potentials; consequently, each fiber will contain a similar equivalent source density. This is shown as a dipole density and hence proportional to -Vm /x. Note that in the aforementioned region the function -Vm (x)/x is monophasic, and hence the dipole sources are all oriented in the same direction.
When the extent of the rising phase of the action potential (x2 - x1 in Figure 8.4) is small compared to the distance to the field point P, then the axial dipole distribution in a small lateral cross-section can be replaced by a lumped dipole. In this case, the source arising in the bundle as a whole can be approximated as a dipole sheet, or double layer. For cardiac muscle, because cells are highly interconnected, the fiber bundle of Figure 8.4 is a good approximation to the behavior of a propagating wave in any cardiac muscle region regardless of the actual physical fiber orientation.
Measurements on laboratory animals permit the determination at successive instants of time of the surface marking the furthest advance of propagation. Based upon the foregoing, these isochronal surfaces may also be viewed, at each instant, as the site of double layer source. Since the thickness of the rising phase of the propagated cardiac action pulse is only around 0.5 mm (1 ms rise time multiplied by 50 cm/s propagation velocity), the condition that it be small as compared to the distance to the field point is nearly always satisfied when considering electrocardiographic voltages at the body surface. The double layer source model is considered by many to be fundamental to electrocardiography..
0) are assumed to be planar. All fibers in the bundle are assumed to be parallel and carrying similar action potentials; consequently, each fiber will contain a similar equivalent source density. This is shown as a dipole density and hence proportional to -Vm /x. Note that in the aforementioned region the function -Vm (x)/x is monophasic, and hence the dipole sources are all oriented in the same direction.
When the extent of the rising phase of the action potential (x2 - x1 in Figure 8.4) is small compared to the distance to the field point P, then the axial dipole distribution in a small lateral cross-section can be replaced by a lumped dipole. In this case, the source arising in the bundle as a whole can be approximated as a dipole sheet, or double layer. For cardiac muscle, because cells are highly interconnected, the fiber bundle of Figure 8.4 is a good approximation to the behavior of a propagating wave in any cardiac muscle region regardless of the actual physical fiber orientation.
Measurements on laboratory animals permit the determination at successive instants of time of the surface marking the furthest advance of propagation. Based upon the foregoing, these isochronal surfaces may also be viewed, at each instant, as the site of double layer source. Since the thickness of the rising phase of the propagated cardiac action pulse is only around 0.5 mm (1 ms rise time multiplied by 50 cm/s propagation velocity), the condition that it be small as compared to the distance to the field point is nearly always satisfied when considering electrocardiographic voltages at the body surface. The double layer source model is considered by many to be fundamental to electrocardiography..
Vm/x, which is shown. A physical representation of this dipole distribution is also shown. The dipoles lie in the range x1 < x < x2.
The source-field relationship for an isolated fiber is described by Equation 8.17, which identifies the source density as the transmembrane current. It was pointed out that when this expression was obtained, the source was approximated as a point (rather than a ring), and that the effect of the fiber itself within the volume conductor was ignored. For the isolated fiber, where the spatial extent of the nerve impulse is large compared to the fiber radius, it can be shown that the line-source formula of Equation 8.17 is very satisfactory (Trayanova, Henriquez, and Plonsey, 1989).
When these conditions are not satisfied, it is desirable to have a rigorous (exact) source expression. One can show that for an arbitrarily shaped active cell of surface S, the field generated by it at point P, outside or inside the cell, is
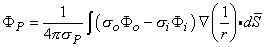 | (8.28) |
| where | Φ P | = field at point P |
| Φ i | = potential just inside the membrane | |
| Φ o | = potential just outside the membrane | |
| σ i | = conductivity inside the membrane | |
| σ o | = conductivity outside the membrane | |
| σ P | = conductivity at the field point |
The source identified by Equation 8.28 is a double layer lying on the membrane surface, whose strength is (σoΦo - σiΦi)dS and whose orientation is along the outward surface normal (Plonsey, 1974). The field point P in Equation 8.28 can be intracellular as well as extracellular; however, the coefficient P takes on the conductivity at the field point.
If one applies Equation 8.28 to an isolated cylindrical fiber, then assuming only Φo 0 (hence Φi - Φo Vm ) leads to
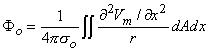 | (8.29) |
where the integration proceeds over the cross-sectional area A, as well as axially. If the field point is at a large distance compared to the radius, then Equation 8.29 reduces to Equation 8.21 and Equation 8.17, thus confirming the earlier work when these approximations are satisfied.
In this section we discuss the mathematical basis of the concepts of volume source density (flow source density), IF, and impressed current density,
8.5 MATHEMATICAL BASIS FOR MACROSCOPIC VOLUME SOURCE DENSITY (FLOW SOURCE DENSITY) AND IMPRESSED CURRENT DENSITY
PRECONDITIONS:
Source: Layer of dipole source elements i
Conductor: Infinite, homogeneous
i.
As a consequence of the activation process in cardiac tissue, the heart behaves as a source of currents and generates potentials in the surrounding volume conductor. These sources consist of layers of dipole source elements, which lie in the isochronal activation surfaces, as pointed out earlier. This description is only an approximation, since it is based on the assumption that cardiac tissue is homogeneous and isotropic.
In principle, Equation 8.28 can be applied to each active cell in the heart. Since a cardiac cell is very small compared to the distance of observation, the radius vector 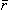 in Equation 8.28 may be assumed to be constant in the integration over each cell. Thus each cell can be thought of as contributing a single lumped dipole source, which is simply the vector sum of its double layer surface elements. That is, the dipole for the jth cell, dj is given by
in Equation 8.28 may be assumed to be constant in the integration over each cell. Thus each cell can be thought of as contributing a single lumped dipole source, which is simply the vector sum of its double layer surface elements. That is, the dipole for the jth cell, dj is given by
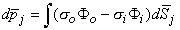 | (8.30) |
Since the heart contains around 5  1010 cells of which perhaps 5% are active at any moment during depolarization, the number of dipole source elements is extremely high. Under these conditions one can define a volume dipole moment density function (i.e., a dipole moment per unit volume) by averaging the dipole elements in each small volume. That is,
1010 cells of which perhaps 5% are active at any moment during depolarization, the number of dipole source elements is extremely high. Under these conditions one can define a volume dipole moment density function (i.e., a dipole moment per unit volume) by averaging the dipole elements in each small volume. That is,
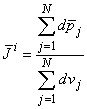 | (8.31) |
where the denominator is the total volume occupied by a group of N cells, and dSj is the surface of each volume element dvj. The idea is to make N small enough so that a good resolution is achieved (where the average is not smoothed unnecessarily), but large enough so that the function i is continuous from point to point (and does not reflect the underlying discrete cellular structure). Equation 8.31 is sometimes described as a coarse-grained average, since we do not let the volume, over which the average is taken, go to zero. The same considerations apply, for example, in electrostatics, where the charge density is normally considered to be a smooth, well-behaved function even though it reflects a discrete collection of finite point sources.
The source function i is a (volume) dipole density function. Consequently, the field it generates can be found by superposition, where idv is a single dipole to which Equation 8.12 applies. Thus, summing the field from all such elements, one obtains
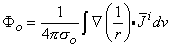 | (8.32) |
If one applies the vector identity  (i /r) = (1/r)i + (1/r)i to Equation 8.32, then
(i /r) = (1/r)i + (1/r)i to Equation 8.32, then
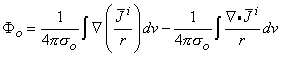 | (8.33) |
The divergence (or Gauss's) theorem can be applied to the first term on the right-hand side of Equation 8.33, and since i = 0 at S (all source elements lie within the heart, and none are at the surface of integration), we get
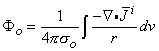 | (8.34) |
Reference to Equation 8.7 identifies that
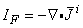 | (8.35) |
is a volume source (flow source) density.
>0:x1 < x < x2
Henriquez CS, Plonsey R (1988): The effect of the extracellular potential on propagation in excitable tissue. Comments Theor. Biol. 1: 47-64.
Plonsey R (1974): The formulation of bioelectric source-field relationships in terms of surface discontinuities. J. Franklin Inst. 297:(5) 317-24.
Plonsey R, Barr RC (1987): Interstitial potentials and their change with depth into cardiac tissue. Biophys. J. 51: 547-55.
Rosenfalck P (1969): Intra- and extracellular potential fields of active nerve and muscle fibers. Acta Physiol. Scand. 321:(Suppl) 1-168.
Trayanova N, Henriquez CS, Plonsey R (1990): Limitations of approximate solutions for computing the extracellular potential of single fibers and bundle equivalents. IEEE Trans. Biomed. Eng. BME-37: 22-35.
As was discussed in Section 7.2.2, one can interpret i as an impressed (i.e., an applied) current density. This current density is brought into being by the expenditure of chemical energy (i.e., the movement of ions due to concentration gradients); it is the primary cause for the establishment of an electric field. In contrast, we note that the current density, = σ , that is described by Ohm's law in Equation 8.4, is induced (i.e., it arises secondary to the presence of the aforementioned electric field ). Impressed currents i are not established by the electric field , since they originate in a source of energy, which is nonelectric in nature.
8.6 SUMMARY OF THE SOURCE-FIELD MODELS
Table 8.1 gives the equations used in this chapter ( with equation numbers) for the different sources and their fields in an infinite homogeneous volume conductor.
Table 8.1. Summary of the equations for different sources and their fields.
Chpt
Source model
Source element
descriptionField Φo outside the source
Source density
8.2
Monopole
Io
(8.7)
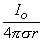
(8.7)
[point source]
Dipole
= Io(8.9)
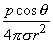
(8.15)
[point (dipole) source]
Single
isolated
fiber r » a
(where a =
fiber radius)im(x)dx
(8.16)
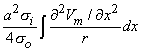
(8.21)
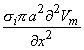
line
source
density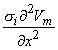
volume
source
density
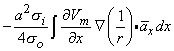
(8.23)
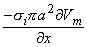
line
source
density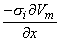
volume
source
density8.3
Fiber bundle
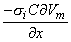
volume
source
density
C 0.4Tripole
σi
2Vm/x2
>0:x2 < x < x3
>0:x3 < x < x4
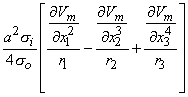
Vm refers here to a triangular
approximation of the actual
transmembrane potential
(exact form of the equation
in the text).(8.27)
Point sources on axis location: strength: x1 
x1 
x1 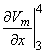
8.4
Single cell
(exact
formulation)(σoΦo - σiΦi)dS
(8.28)
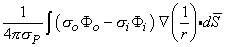
(8.28)
Double layer with
a strength of
(σoΦo - σiΦi) lying
in the cell membrane,
and oriented in the
outward direction
Isolated
cylindrical
fiber (exact
formulation)(σoΦo - σiΦi)dS
(8.28)
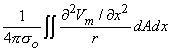
(8.29)
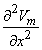
volume source
(flow source)
density
lying within
the fiberMulticellular
tissue
(brain or
cardiac
tissue)idV
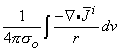
(8.34)
ivolume dipole
density-
ivolume source
(flow source)
densityREFERENCES